
细胞焦亡
5.6k
ZKEASOFT
2020/03/30
焦亡是一种程序性坏死细胞死亡,在存在病原体相关的分子模式(PAMP)或细胞衍生的损伤相关的分子模式(DAMP)的情况下,细菌、病毒、真菌和原生动物的细胞内感染会激活焦亡。它通常在先天免疫系统的细胞(例如单核细胞、巨噬细胞和树突状细胞)中被诱导。焦亡通常是病原体感染后可能触发的主要细胞死亡方式,人们认为当 caspase 酶无法获得时,其他类型的细胞死亡(如坏死性凋亡)会作为继发过程发生。发生焦亡的细胞显示出诸如细胞肿胀、膜起泡、DNA 断裂和最终细胞裂解等特征。但是,细胞核通常保持完整,这与凋亡和坏死性凋亡中观察到的细胞核破坏现象不同。
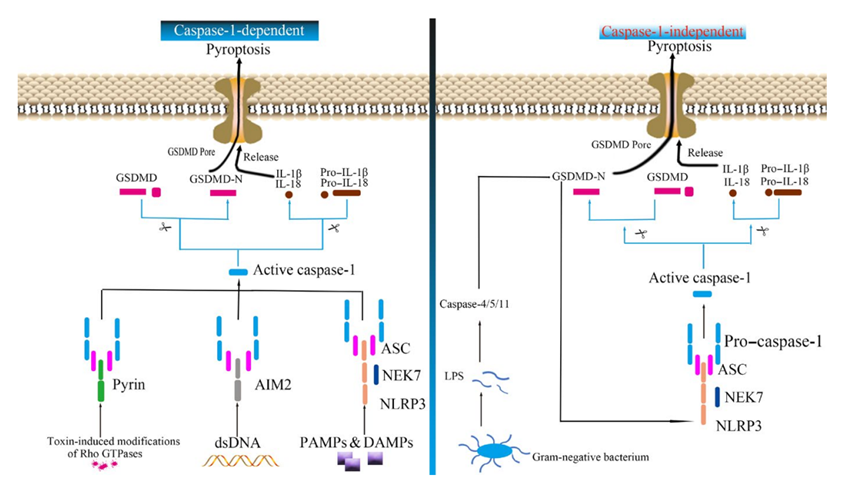
经典通路
焦亡经典通路通常被描述为通过两步过程发生。在第一个“启动信号”步骤中,激活 NF-kB 以诱导多种蛋白质的表达,这些蛋白质将成为所谓的炎性小体的复合体的一部分。炎性小体通常包含一个胞质模式识别受体(PRR;例如 AIM2 或 AIM2 样家族成员)、一个接头蛋白(例如 ASC/TMS1)和促 caspase-1。通过检测 NLRP3 和可视化 ASC 斑点来分析炎性小体的激活是监测焦亡的另一种有用技术。
在第二个激活步骤中,caspase-1 在裂解 gasdermin D 和细胞因子,促 IL-1β 和促 IL-18前被蛋白水解激活,产生其促炎症形式 IL1β 和 IL18,这些是由死亡细胞分泌的。这些重要细胞因子的可视化可以通过使用特异性抗体来实现。重要的是,caspase-1 还可切割关键蛋白 gasdermin D(GSDMD)的氨基末端片段,使其寡聚并在细胞膜上形成孔。这会导致细胞因子分泌、进水和最终的细胞破裂。
非经典通路
另外通过细胞内检测革兰氏阴性细菌 LPS 的非经典通路来诱导焦亡,从而激活 caspase-4、5和/或 11(小鼠)以切割 GSDMD。由于 GSDMD 在经典和非经典通路中均被切割,因此监测 GSDMD 的切割和易位是识别焦亡的重要部分。
疾病研究中的焦亡
研究焦亡的机制可能会为癌症、自身免疫性疾病、神经退行性疾病等带来治疗益处。有研究证明 ω-3 脂肪酸在三阴性乳腺癌细胞中能激活焦亡,表明焦亡具有介导癌症的潜力;然而其机制仍有待阐明。在炎性肠病中,炎症小体激活和 caspase 诱导表明存在焦亡,这可能也是开发新型治疗药物的有前景的研究方向。
细胞焦亡的研究方法
(1) GSDMD的切割 (Western检测);
(2) Caspase的激活,主要是Caspase-1,Caspase-4,Caspase-5,Caspase-11。(Western检测);
(3) IL-1β和IL-18前体的切割成熟和释放 (Western,ELISA等);
(4)细胞形态学检测(CCK-8等);
(5)染色质完整性检测 (Tunel等)。